Cellular Pathways
AO
AbdulSamad Olagunju / November 29, 2021
9 min read
Welcome to another blog post!
Quote of the Post:
"Every life is different from any that has gone before it, and so is every death. The uniqueness of each of us extends even to the way we die." - Sherwin Nuland
These are some important cellular pathways I learned in my Cellular Biology class this past semester. In this blog you can look at some notes I made about these cellular pathways. If you want me to elaborate on any of the pathways in a separate article, feel free to contact me. I may choose to go into more detail in future blog posts.
- Firstly, it will be important to define GEF’s and GAP’s. Take a look at this diagram:
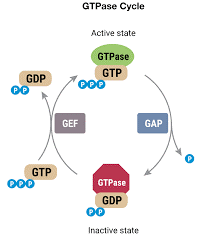
- Image Link
- There are small molecules called GTPases that are very important for regulating cellular pathways.
- The GTPases can be turned on or off. They are off when they are bound to a GDP (guanine diphosphate) and they are on when they are bound to a GTP (guanine triphosphate). Take a look at the picture below and try to figure which molecule is the GTP or GDP (hint: search up what a phosphate is).
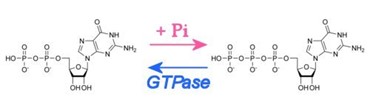
- A GAP will remove a phosphate from GTP, resulting in the GDP being attached to the GTPase.
- A GEF will replace the GDP on the GTPase with a GTP, activating the GTPase.
- GPCR: G Protein Coupled Receptor
- cAMP: cyclic AMP (adenosine monophosphate)
- G Protein:
- G alpha bound to GTP, Ligand binds receptor, G alpha releases GDP and grabs GTP; GEF(Guanine nucleotide Exchange Factor => this activates the GTPase, replace the GDP with a GTP) is GPCR, G protein activated, G alpha interacts with adenylyl cyclase, adenylyl cyclase makes cAMP, GAP (GTPase Activating Protein => this removes a phosphate from the GTP, resulting in GDP) turns GTP to GDP on G alpha
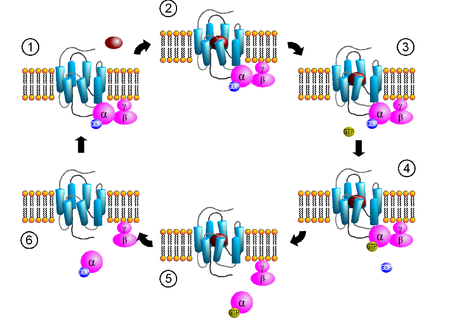
- cAMP turns on Protein kinase A (usually floating in cytoplasm), Protein Kinase A changes shape after cAMP binds to regulatory subunit, kinase phosphorylates serine, threonine, or tyrosine
- G beta gamma can bind to K+ ion channel
- cAMP degraded by phosphodiesterase
- PKA can phosphorylate GPCR and deactivate it, deactivates GEF function of GPCR
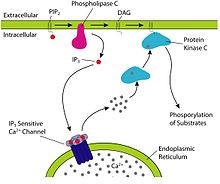
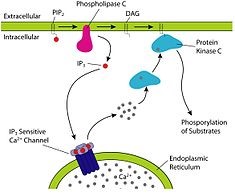
- IP3
- Ligand binds to GPCR, G protein activated, Phospholipase C cuts PIP2 into IP3 and DAG, IP3 opens Ca2+ channel, DAG activates protein kinase C
- Receptor Tyrosine Kinase (RTK):
- Ligand binding module and tyrosine module on RTK, ligand binding activates tyrosine kinase, tyrosine kinase only phosphorylates tyrosine residues, autophosphorylation
- Proteins with src domain (kinase domain, sh2 and sh3 domain, sh2 binds to phosphorylated tyrosine), c terminus is phosphorylated or its not, when c terminus is not phosphorylated, src kinase domain can phosphorylate stuff
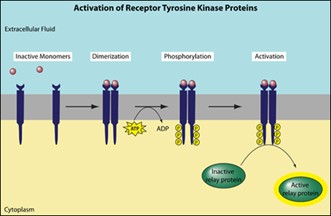
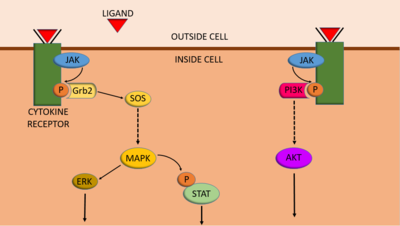
- JAK/STAT:
- RTK dimerization due to ligand, kinase is separate polypeptide clinging to cytoplasmic domain of receptor, kinase then activates STAT
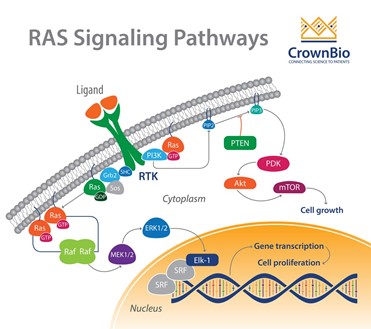
- Ras Signaling (Important for Growth factors, cancer => hyperactivated Ras):
- Ras signaling is extremely important for growth factors, and mutations to the Ras protein are often implicated in cancer. Let us try to understand Ras signaling.
- Activate RTK
- Sos floating in cytoplasm, bound to GRB2
- GRB2 has sh2 domain, binds to phosphorylated tyrosine
- GRB2 releases Sos
- GEF called Sos activated
- Sos acts as a GEF for Ras, Ras (member of Arf, Sar family) is activated and replaces GDP with GTP
- Ras interacts with Raf, Raf kinase then activated
- Raf activates MEK
- MEK activates MAP kinase
- MAP kinase activated
- MAP kinase goes in nucleus, phosphorylates Ets and Jun
- Ets and Jun turn on genes (transcription factor)
- Ras inactivated by GAP, GDP bound Ras is inactive
- Phosphatase can reverse steps in pathway
- Image Link
- Phospholipase C (has sh2 domain):
- Activated by RTK
- Leads to production of DAG and IP3
- Yeast
- Mating factor activates G protein, G beta and gamma bind to scaffolding protein
- Ste5 (Ste20, MEKK, MEK, MAPK)
- Mating pathway genes expressed
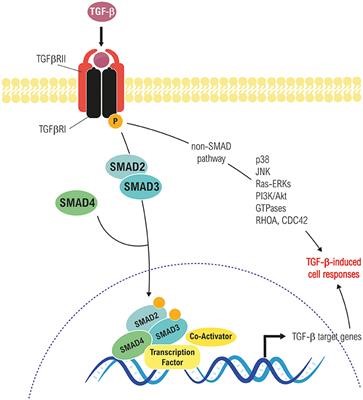
- TGF Beta
- Tetramer forms between Type 1 and 2 receptor
- Type 2 phosphorylates type 1
- R-smad phosphorylated
- R-smad interacts with smad4
- Smad complex enters nucleus, acts as transcription factor
- Link to Image
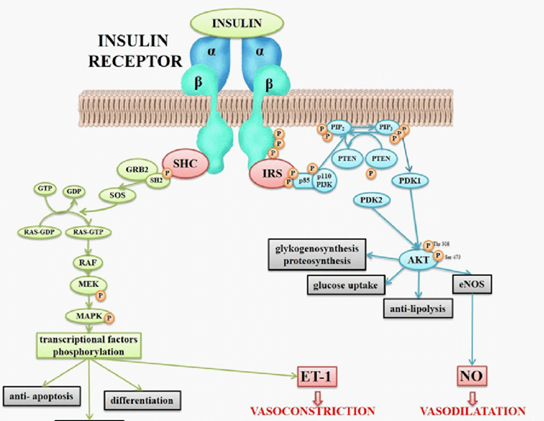
- Insulin
- Insulin receptor is always a dimer
- Insulin binds, conformational change that activates kinase, autophosphorylation
- IRS-1 protein phosphorylated
- IRS-1 can activate Ras-MAPK pathway (Interacts with GRB2)
- IRS-1 can activate PI 3-kinase
- PIP2 is substrate for PI 3-kinase (PIP2 is also substrate for Phospholipase C)
- PI 3-kinase phosphorylates PIP2 to PIP3
- PTEN is phosphatase, can turn PIP3 to PIP2
- PIP3 binds to Akt
- Akt activated, Akt phosphorylates proteins, leads to GLUT4 being made
- Link to Image
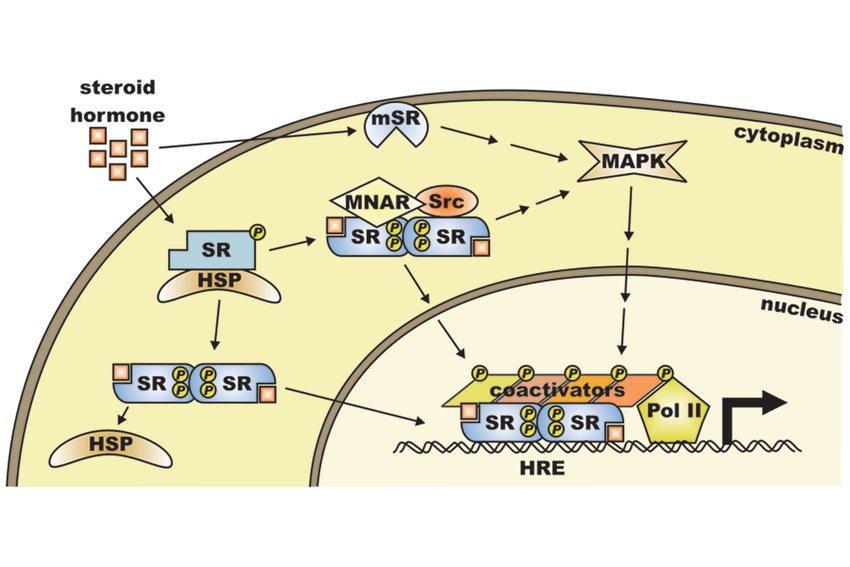
- Steroid binds to Hsp-Receptor complex, Hsp protein leaves, steroid complex enters nucleus
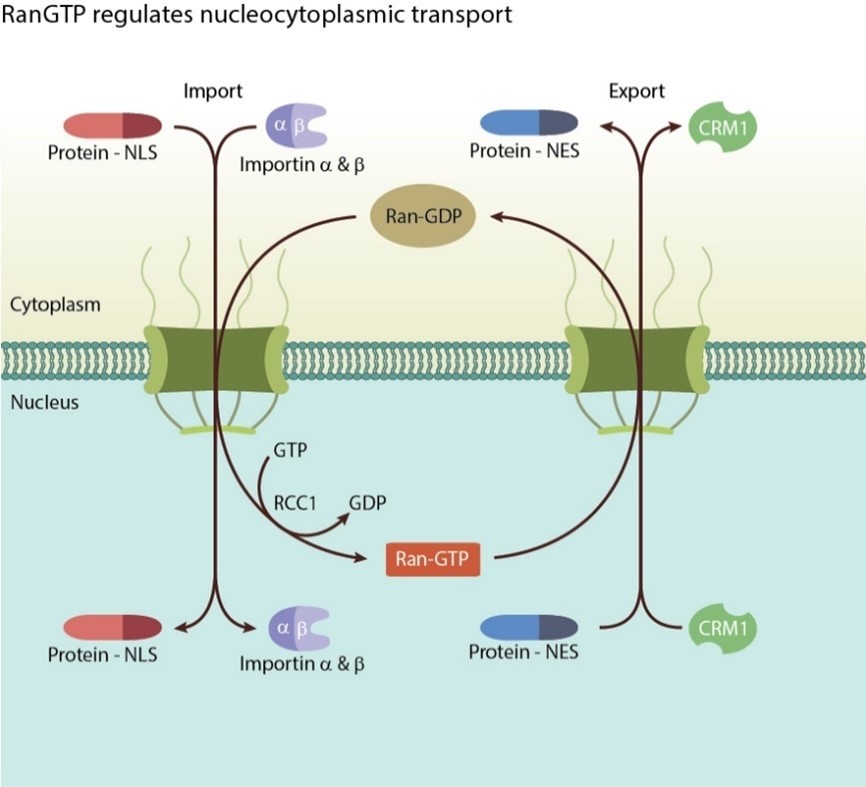
- Import Process:
- Protein made in cytoplasm with nuclear localization sequence, importin forms complex with protein, diffuses through nuclear pore (unstructured proteins in nuclear pore), Ran GDP hit by GEF (GEF for Ran in nucleus), RanGTP causes importin to release cargo, RanGTP importin complex diffuses out of nucleus, Ran GAP in cytoplasm causes RanGTP to turn to RanGDP and release importin
- Ran GDP shuttled back to nucleus
- Link to Image
- Export process:
- mRNA needs to get out, forms mRNA/protein complex, has nuclear export sequence; tripartite complex between RanGTP, mRNA, and exportin; move out of nucleus to cytosol, GAP causes Exportin and cargo to be released
- Ran GDP shuttled back to nucleus
- Link to Image
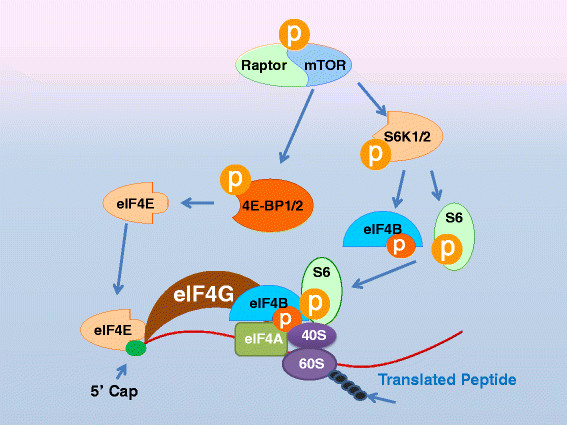
- TOR Kinase:
- Activated by growth factor
- Nutrient sensor
- Cap dependent translation:
- TOR Kinase phosphorylates 4E-BP1 (so can EK/MAPK, PKA, PKC)
- 4E-BP1 job is to inhibit cap dependent translation protein (eIF-4E)
- Cap dependent translation can proceed
- Link to Image
- 5’ TOP mRNA
- Not every mRNA has 5’ cap, 5’ TOP mRNA’s have different cap, they only encode things needed for cell division
- TOR phosphorylates S6K
- S6K phosphorylates ribosomal protein, allows ribosomes to interact with 5’ TOP mRNA
- When you need to make way more ribosomes, use 5’ TOP mRNA’s
- Link to Image
- Cyclin dependent kinases:
- Kinase that needs to be in complex with cyclin
- Need to activate CDK to progress through checkpoint, cell needs to produce cyclin to progress through checkpoint
- Wee1 adds two inhibitory phosphorylations to (cyclin dependent kinase bound to cyclin)
- Cyclin Activating Cyclin (CAK) has activating phosphorylation
- Cdc25 (cell division cycle 25) removes the 2 Wee1 phosphorylations
- CDK phosphorylates Cdc25 to increase its activity (Cdc25 is substrate for activated CDK)
- Anaphase-Promoting Complex is also substrate of CDK
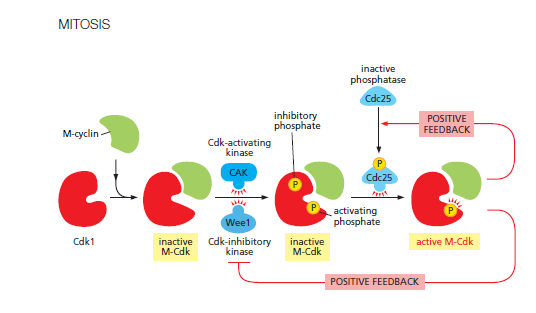
- Link to Image
- Anaphase promoting complex (APC)
- Needs Cdc20
- CDK phosphorylates APC
- APC phosphorylated, triggers recruitment of cdc20
- Cdc20 only made after CDK activated
- Activated APC degrades cyclin, shuts off CDK
- APC targets securin
- Cohesins hold together sister chromatids
- Separase inhibited by securin
- APC destroys securing, separase activated, cohesin cleaved
- Mad and Bub (part of kinetochore complex)
- Make sure K fibers attached to chromosome
- Inactivate cdc20 so that it can’t join APC
- When chromosomes attached to K fibers, cdc20 released and Mad and Bub inactivated
- Licensing occurs in G1, ORC complex (origin recognition complex), bind to origin of replication, helicase loader recruited (cdc6 and cdt1), MCM proteins then recruited (including helicases, helicases use lots of ATP)
- CDK pushes from 1 phase to the next, prevent origins from being licensed again in S phase by CDK phosphorylating ORC proteins and helicase loaders, trigger degradation of some proteins
- Geminin: inhibitor of licensing, made in G1 (during licensing), Geminin binds to Cdt1 when you exit M phase, geminin destroyed
- P53:
- Constantly made, immediately destroyed in normal cells
- DNA damage detected by cell
- ATM and ATR activated (they are kinases)
- ATM and ATR phosphorylate p53
- Mdm2 (E3 ligase) can’t ubiquinate p53 and destroy it when it is phosphorylated
- p53 can act as transcription factor
- p53 can trigger production of p21 to arrest cell cycle
- p53 can trigger production of Puma
- p21 binds to G1 CDK-cyclin complex, inhibits it; fixable cell damage
- G1 CDK-cyclin complex can’t phosphorylate Retinoblastoma
- Retinoblastoma can bind to E2F1 transcription factor
- E2F1 can’t turn on genes necessary for S phase
- Usually, G1 CDK-cyclin complex phosphorylates Retinoblastoma, Retinoblastoma releases E2F1, S phase happens
- Arrest cell cycle till DNA damage gone, ATM and ATR turn off, p53 not phosphorylated and destroyed
- Puma (DNA damage is too much)
- Puma inhibits Bcl-2
- Bcl-2 inhibits proteins that trigger apoptosis, but it is inhibited
- Apoptosis occurs
- Apoptosis
- In mitochondria, double membrane
- Proteins call BAK and BAX
- BAK and BAX can form into pore
- If pore forms, Cytochrome C, Smac, Diablo, released into cell, which triggers apoptosome
- Bcl-2 holds back pore formation
- If Puma inhibits Bcl-2, mitochondrial pore forms, cell dies
- Caspases are proteases, usually exist in pro (like procollagen) form, if they are activated, they clip stuff
- When caspases clustered together, they clip each other, get mature caspase
- Caspase 8 and 9 are initiator caspases
- Also executioner caspases
- Extrinsic Pathway:
- Ligand interacts with death receptor
- Activate caspase 8 by caspase 8 clustering
- Caspase 8 activates 3, 6, and 7
- Leads to apoptosis
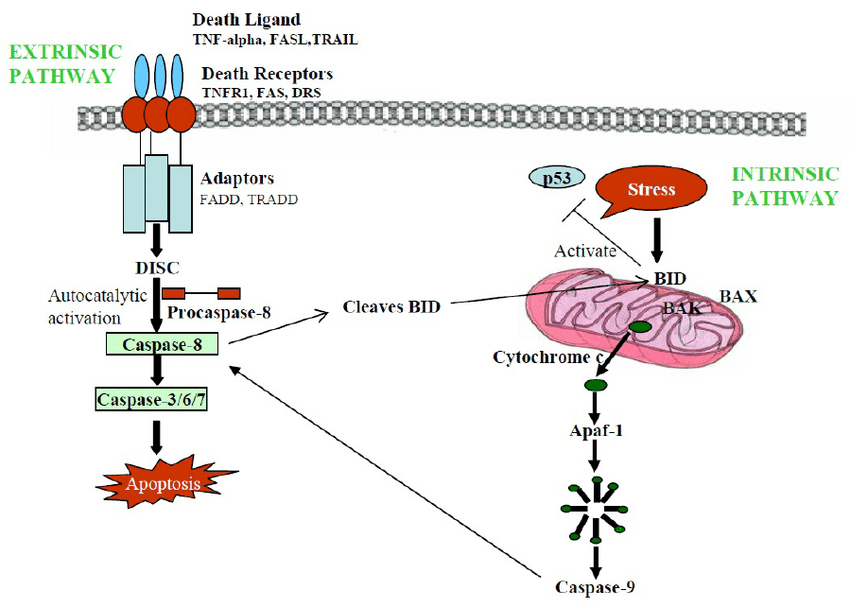
- Link to Image
- Intrinsic Pathway:
- Some sort of cell damage, p53 activated
- Then mitochondria releases cytochrome c
- Cytochrome c scaffold assembly of caspase 9
- Caspases cluster, clip each other
- Caspase 3, 6, 7 activated
- Leads to apoptosis
- Link to Image
Hopefully this helps you with your molecular biology.